The cells are orientation selective and spatially organized according
to a two-dimensional model orientation preference map
[37,61]; see Fig. 10.
In addition to the center-surround connectivity (discussed above),
each cell has 10 long-range connections, chosen in the range between
25-50 lattice constants which connect it only with cells of similar
orientation preference (difference in preferred orientation less than
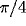 ) reproducing the pattern of long range clustered connections
found in physiological studies
[14,28,29]. In these
simulations we assume that the long-range connections between cells
with similar orientations are excitatory (but see the following
section).
) reproducing the pattern of long range clustered connections
found in physiological studies
[14,28,29]. In these
simulations we assume that the long-range connections between cells
with similar orientations are excitatory (but see the following
section).
 |
 |
| |
|
 cells where the gray-level of each pixel corresponds to
the preferred orientation of the respective cell (lightest gray:
horizontal, light gray:
cells where the gray-level of each pixel corresponds to
the preferred orientation of the respective cell (lightest gray:
horizontal, light gray: 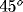 , dark gray:
, dark gray: 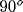 , black:
, black: 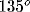 ).
White pixels represent spiking cells. At this time, most of the
activity is in regions with preferred orientation of about
).
White pixels represent spiking cells. At this time, most of the
activity is in regions with preferred orientation of about 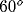 .
.
External input is modeled independently for each cell as a Poisson
process of excitatory voltage pulses of size 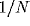 , arriving at a mean
rate which depends on the phase difference between the stimulus
orientation and the cell's preferred orientation. The simulations were
performed under two conditions:
, arriving at a mean
rate which depends on the phase difference between the stimulus
orientation and the cell's preferred orientation. The simulations were
performed under two conditions:
In particular, we compare the strength of synchronization in the stimulus condition between cells that receive the same stimulus (both in the right half plane or both in the left half plane) versus cells which receive mutually orthogonal stimuli. Figure 10, left shows a snapshot of the activity in the 2-dimensional array, superimposed on the underlying orientation column structure.
For mutually excitatory pairs of cells, the cross-correlations show a `castle' [35] with a width of about 10-20 msec and centered at zero time offset (Fig. 5, top right). The cross-correlation function between model neurons at larger distance shows a weaker, slower ``hill'' (Fig. 5, bottom right). For mutually inhibitory cells the correlation function is characterized by a dip that rises to the coincidence level at longer times (Fig. 5, center right).
 |
 |
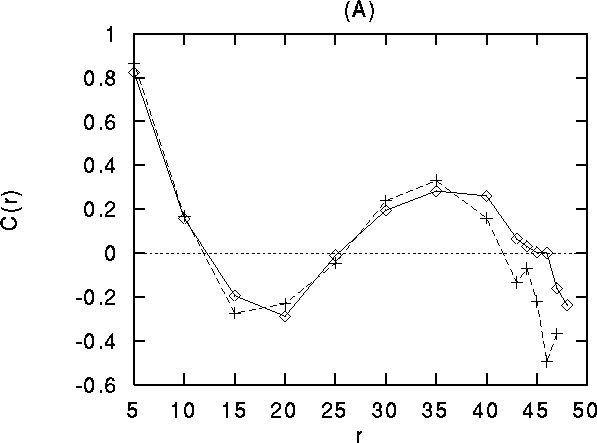
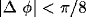 ), crosses: between cells of orthogonal orientation
(
), crosses: between cells of orthogonal orientation
(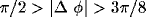 ;
; 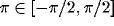 ). For
distances larger than about 35 lattice constants, correlations
between cells with common orientations (diamonds) become stronger than
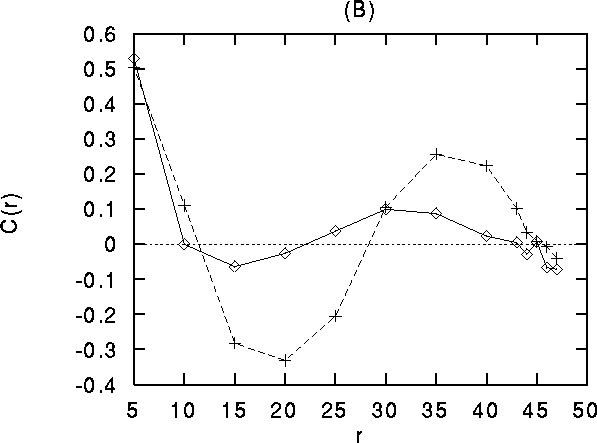
those between orthogonal orientations. (B) Differential stimulation.
Diamonds: correlation coefficient between cells receiving stimulation
with different orientation, crosses: correlation coefficient between
cells receiving stimulation with the same orientation. Both curves in
(B) are averaged over cells with all orientations. Note that for r
between 30 and 45 (about the range of long-distance connections),
pairs of cells receiving input with similar orientation are more
strongly correlated than cells receiving input with orthogonal
orientations, even if the members of the pair have with different
preferred orientation.
). For
distances larger than about 35 lattice constants, correlations
between cells with common orientations (diamonds) become stronger than
those between orthogonal orientations. (B) Differential stimulation.
Diamonds: correlation coefficient between cells receiving stimulation
with different orientation, crosses: correlation coefficient between
cells receiving stimulation with the same orientation. Both curves in
(B) are averaged over cells with all orientations. Note that for r
between 30 and 45 (about the range of long-distance connections),
pairs of cells receiving input with similar orientation are more
strongly correlated than cells receiving input with orthogonal
orientations, even if the members of the pair have with different
preferred orientation.
We quantify the strength of synchronization by the contribution coefficient of the cross-correlation [51]. This quantity measures the number of coincidences above chance level and is computed as the area of the cross-correlation above the asymptote divided by the total number of spikes. Both in the control case as well as in the simulation with oriented external input (Fig. 11), the contribution coefficient exhibits a spatial oscillation, with largest values at small distances (for neighboring cells), then becoming negative at about 15-30 lattice units (indicating anti-correlated activity), and then becoming positive again at the typical distances of the long range clustered connections (35-45 lattice constants).
In the control situation, the contribution coefficient at large distances is larger for similar vs. orthogonal orientations. For the 2-orientation stimuli, the contribution coefficient of cells with the same separation (30-45) is larger for cells belonging to the same half plane than for cells across the boundary between the two half s of the system that receive different stimuli (even if the later have the same orientation preference).
We have also computed the effect of stimulus-dependent synchronization on the basis of the LFP variables. We have computed the mean LFP cross-correlations between two recording sites, stimulated either by the same or by different gratings, within the same distance between each pair of sites. As shown in Figure 12, the correlation between cells receiving the same input is significantly stronger than that between cells that receive different inputs. One should notice that this effect is stimulus dependent and can not be explained by the simple fact that connected cells synchronize [31]. In fact, the degree of correlation between two given cells depends their stimulation and is not only a function of the connectivity.
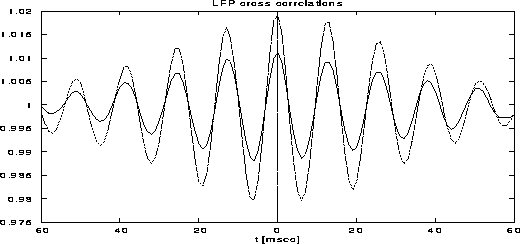
Figure 12:
Cross-correlation functions of local field potentials.
The dotted curve is between populations of cells with the same preferred
orientation and the full line is between different preferred
orientations.
The mechanism responsible for stimulus dependent synchronization is the following:
Thus we find stimulus-dependent synchronization for cells in the range of the clustered connections. In accordance with recent experimental findings [16] and with other modeling work [45], we find that the degree of synchronization is higher when cells are excited with the same orientation. Synchronization is strong between nearby cells (independent of orientation preference) and there is a clear tendency for synchronization between cells with the same orientation preference separated by larger distances. We find that this result does not require models based on abstract phase oscillators with amplitude-dependent interactions, as in [45], but can be obtained in the more biologically plausible framework of populations of integrate-and-fire neurons.