In the simulations presented in the previous sections we have assumed that the long range connections between cells with the same orientation are excitatory. In reality, the nature of these long-range connections is more complex, revealing both excitatory and inhibitory influences [20]. For example, modulatory influences on the response of a cell from outside its ``classical'' receptive field (CRF) have been observed in a number of physiological experiments for both moving and stationary stimuli, both in cats and primates [17,23,34]. In most cases, presentation of extended arrays of similar stimuli leads to a partial suppression of the cell response; response increases were observed less frequently. Furthermore, the suppression is larger if line segments outside the CRF have the same orientation as the segments inside the CRF. A straightforward explanation of these results in terms of a weak inhibitory influence of the surround elements is, however, incompatible with other experimental results [17,23] that show that presentation of the same surround stimulus alone leads to a weak excitation of the center cell.
We have recently addressed the balance between excitation and inhibition in the long-range horizontal connections [47] within the context of the network model described above [47]. To simplify matters, we assumed that the CRF maps onto a single hypercolumn in the V1 map of orientation preferences, while surround stimuli excite cells in neighboring hypercolumns. Given the different characteristics of excitatory and inhibitory cells, the response to additional input through the lateral connections depends on the firing rate of the local population responding to the CRF stimulus. More explicitly, for very weak (sub-threshold) `center' stimulation , a surround stimulation of the same orientation as the center stimulation is mainly excitatory, while for a strong center stimulus the surround contributes mainly inhibition (see Figure 13), explaining thus the experimental data reported above.
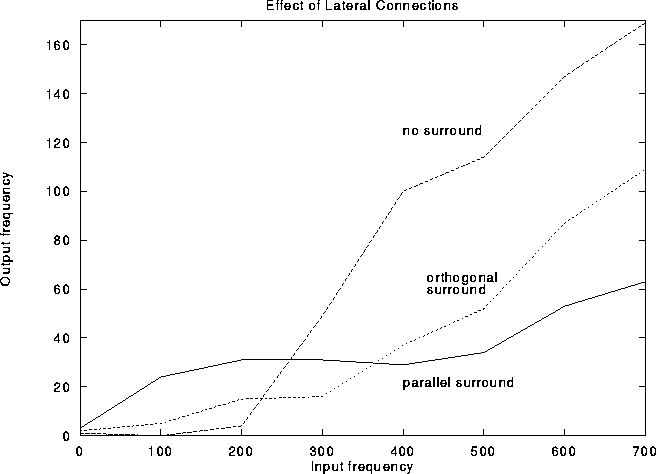
Figure 13: Average response curves (``F-I curves'') of cells within
the classical receptive field as a function of LGN
input frequency. The central stimulus is always at the
preferred orientation. The dashed curve is in the
absence of any additional input. The dotted and full lines were
obtained by adding orthogonal and parallel surround stimuli,
respectively. Note that all surround input enhances the
response frequency for low input and suppresses the response for large
center input. Note, furthermore, that surround input always suppresses
large inputs but more so for same-orientation surround input than for
orthogonal surround input.
As we further discuss in [47], this dual effect is a result of the stochastic nature of the spiking input to cells (stochastic resonance) and of the differential excitability of pyramidal and stellate cells and inhibitory neurons.
The computational consequence of activity-dependent modulation is obvious. An efficient information processing system should enhance local differences by suppressing the response to homogeneous stimuli, thereby reducing input redundancy. Such effects may correspond to psychophysical observations of pop-out [5] and tilt-illusion phenomena [59] and seem to require inhibition between cells with similar orientation preferences. On the other hand, when the system is presented with degraded information or weak stimuli, the most efficient strategy consists in performing correction processes such as line completion and stimulus enhancement. Indeed, [40,41] showed that the detection threshold of an oriented stimulus is lowered by the presence of collinear ``flanking'' stimuli with the same orientation. Such an effect is consistent with excitation between orientation columns tuned to similar orientations (modulating a weak stimulus). As shown in Figure 14, simulations show that the excitatory surround contribution (obtained when the center-stimulation is weak) indeed leads to a decrease in the detection threshold.
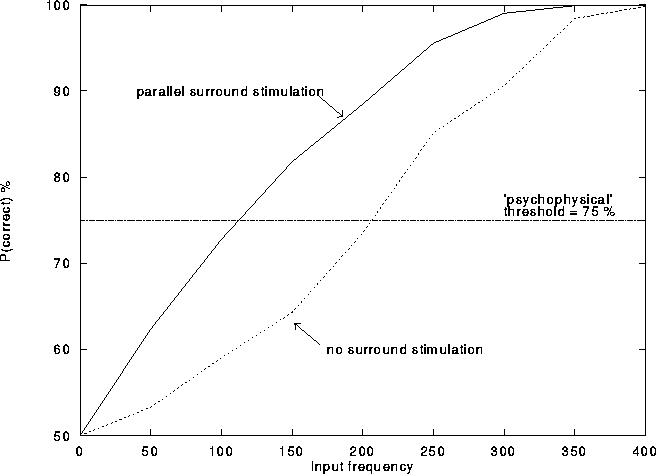
Figure 14:
The addition of parallel surround stimulation lowers
the detection threshold of the center stimulus at weak input intensities.
300 trials were run under each stimulus condition; the spike count
of one cell within 150 msec of stimulus onset was recorded for each
trial. For each surround stimulus condition, the curve represents
the probability, given any two trials, one with a stimulus,
and one without, that the trial with the stimulus produced
more spikes than the trial without a central stimulus.