
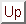

Dynamic receptive fields are observed in response to temporary artificial scotomas in the retina. Such a lesion prevents part of the cortex from receiving input, producing a corresponding cortical scotoma. However, if the surround of the scotoma is stimulated for a period of several minutes, and the scotoma is then removed, the receptive fields of the unstimulated neurons are found to have expanded [35]. The expansion is largest along the preferred orientation of each neuron. Psychophysical experiments further show that after removing the scotoma, a stimulus at the edge of the scotoma appears to have shifted towards the center [24]. Prima facie, such dynamic expansion of receptive fields, and the perceptual shift accompanying it, is incompatible with Hebbian self-organization (which are based on coincident activity), and has been difficult to explain.
 |
 |
| |
|
The laterally connected self-organizing model suggests an explanation. Figure 5 shows how the topographic map reorganizes when a retinal scotoma is introduced and inputs are presented in the surrounding area. The receptive fields of the central, unstimulated neurons remain in the same location as before. Neurons that have part of their receptive fields outside the scotoma are stimulated by the surrounding input, and by Hebbian adaptation, reorganize their afferent weights into the periphery of the scotoma. As a result, these neurons become insensitive to the central region. If the scotoma is now removed, and an input is presented in the scotoma region, only the previously unresponsive neurons respond vigorously to the new input; the surrounding ones do not. Therefore, there is considerably less lateral inhibition from the surrounding neurons to the central neurons, and responses that were previously suppressed by lateral inhibition are unmasked. Therefore, when the RF sizes of the central neurons are measured (based on network activity), they appear to have increased. The expansion is greatest along the preferred orientation because the strongest afferent weights lie in this direction (figure 2 a, b), and any decrease of inhibition unmasks responses mainly in that direction.
Such a reorganization can account for the psychophysical experiments as well. The neurons whose receptive fields have moved outward now respond to inputs farther from the center than before. Therefore, an input at the edge of the retinal scotoma stimulates many neurons inside the cortical scotoma that previously would not have responded, and the response pattern is shifted inward, producing the perceptual shift. After the scotoma is removed and the normal stimulation reestablished, the reorganized RFs gradually return to the normal state (of figure 5 a), and the shift disappears. The model thus shows how the same self-organizing processes and lateral interactions that sculpt the receptive fields during early development could, in the adult, maintain them in a continuously adapting, dynamic equilibrium with the visual environment.