

We have examined experimentally the key prediction of the model, that the same surround stimulus has opposite effects on cell responses depending on the level of excitatory drive from the center. We imaged intrinsic signals in V1 of adult cats in order to examine responses of neuronal populations (see methods). The experimental stimuli used represented the four extremes of the contrast ranges tested in the model: (a) no surround, with center at zero contrast (``blank'') and at high contrast (a ``center'' stimulus), and (b) high-contrast surround, with center at zero contrast (a ``surround'' stimulus) and at high contrast equal to that of the surround (a ``full-field'' stimulus). These stimuli are shown schematically in figure 5, except for the ``blank'', which is used as the baseline reference for the other images. Surround gratings were always presented at the same orientation, velocity and phase as the centers. Orientation was varied systematically, and maps of orientation preference were obtained for each stimulus condition (see methods).
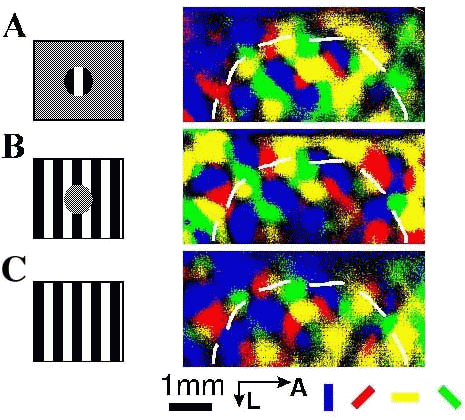
Figure 5:
Optical imaging of intrinsic signals in cat V1 in response to
(A) ``center'', (B) ``surround'', and (C) ``full-field'' stimuli.
The visual
stimulus is depicted in the left column (actual size of center spot,
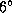 ).
Four orientations of each grating were presented, and 50 trials for
each orientation were summed. The right column shows a map of orientation
domains in which pixel color is used to code the angle of preferred orientation
(color key shown beneath figure) and intensity to code the magnitude of
orientation-selective response. Images were clipped equally. The nominal
retinotopic representation of the center/surround edge was determined from
``center'' and ``surround'' images as the border where activity was equal in
the two stimulus conditions, and is shown on the composite maps in the
right panel as a white dashed line. Extensive activity occurred across
this border in both ``center'' and ``surround'' maps, though in an
orientation-specific manner. A, anterior; L, lateral.
).
Four orientations of each grating were presented, and 50 trials for
each orientation were summed. The right column shows a map of orientation
domains in which pixel color is used to code the angle of preferred orientation
(color key shown beneath figure) and intensity to code the magnitude of
orientation-selective response. Images were clipped equally. The nominal
retinotopic representation of the center/surround edge was determined from
``center'' and ``surround'' images as the border where activity was equal in
the two stimulus conditions, and is shown on the composite maps in the
right panel as a white dashed line. Extensive activity occurred across
this border in both ``center'' and ``surround'' maps, though in an
orientation-specific manner. A, anterior; L, lateral.
Figure 5 shows the cortical images in response to these stimuli in one animal. The ``center'' stimulus elicited several foci of activity (figure 5A) whose location varied with grating orientation. A composite map of the activity at all orientations was consistent with the map of orientation preference described previously in V1 (area 17) [33] and in V2 (area 18) of cats [5]. Interestingly, activity extended over a large region of cortex, to include regions where classical receptive fields of single neurons did not overlap the stimulus (see below). The ``surround'' stimulus elicited orientation-specific activity from a large cortical region as well (figure 5B), including nearly the entire central portion of cortex activated strongly by the ``center'' stimulus. The surround thus ``filled in'' the central cortical representation. When compared to the ``blank'' stimulus, adding the surround increased the activation of the central cortex. The "full-field" stimulus also elicited activity over the entire imaged cortex (figure 5C). However, the magnitude of activation within central cortex was less during ``full-field'' stimulation compared to ``center'' stimulation (figure 5A,C, right panel).
To measure specifically the effect of extraclassical receptive field stimulation on a population of neurons, we recorded single unit receptive fields from the central and peripheral regions of the imaged cortex. The outline of the ``center'' stimulus is superimposed on the fields recorded within the center representation. For neurons at this location, the ``surround'' stimulus engaged primarily the extraclassical receptive field. From such combined single unit and optical imaging experiments (n=4 animals), and in agreement with previous receptive field studies [2], it was clear that at distances >2 mm from the center/surround border (see legend to figure 5), the surround stimulus exclusively engaged the extraclassical receptive fields of neurons. At such discrete locations well within the center representation, we calculated the level of activity for the stimulus conditions above.
We selected for analysis a small region (roughly 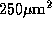 ) well
within the ``center'' region of the map (see text), and avoiding pinwheel
centers. The vector magnitude (independent of vector angle) in this region
was averaged and compared across the different stimulus conditions. We
found the vector magnitude of a pixel to be a more reliable indicator of
signal strength than the image intensity itself. This had the advantage
of being relatively immune to small fluctuations in the gain of the data
collection system.
For robustness of magnitude estimation, images from
at least 8 stimulus orientations were obtained. The analysis is equivalent
to obtaining pixel by pixel the magnitude of the second Fourier component
across angle space
[39].
The strength of the optical signal appears to correspond
well to the magnitude of neuronal activity in cortex
(see also
[12]).
) well
within the ``center'' region of the map (see text), and avoiding pinwheel
centers. The vector magnitude (independent of vector angle) in this region
was averaged and compared across the different stimulus conditions. We
found the vector magnitude of a pixel to be a more reliable indicator of
signal strength than the image intensity itself. This had the advantage
of being relatively immune to small fluctuations in the gain of the data
collection system.
For robustness of magnitude estimation, images from
at least 8 stimulus orientations were obtained. The analysis is equivalent
to obtaining pixel by pixel the magnitude of the second Fourier component
across angle space
[39].
The strength of the optical signal appears to correspond
well to the magnitude of neuronal activity in cortex
(see also
[12]).
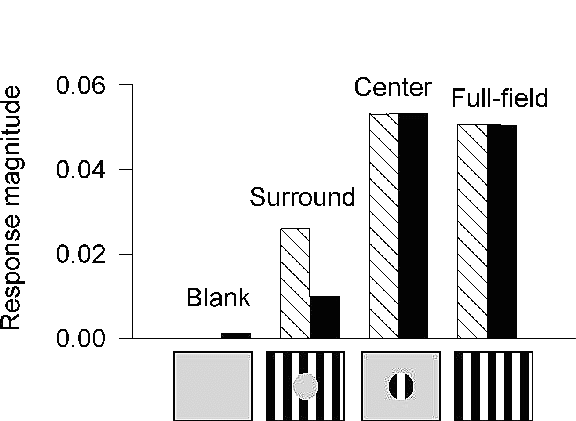
Figure 6: Dynamic modulation of responses
in V1, as shown by the model and optical imaging experiments.
The experimental data (hatched bars) shows a comparison of the intrinsic
signal generated by each of the visual stimuli in a small patch of cortex
within the ``center'' region of the optical map . We determined by single unit
recording that the ``surround'' stimulus exclusively
engaged the extraclassical receptive fields of neurons in this patch. The
average magnitude of the orientation vector is shown as a percentage of the
collected signal. The model's activity measure (black bars) is the
sum of all spikes and the scaled sum of all PSP conductances (factor=1000)
for 180 cells (6x6 columns) in the central-most iso-orientation domain. For
purposes of comparison, the model and imaging data were normalized such that
the center responses were equal. Both the model and experiment demonstrate
that ``surround'' stimulation in the absence of center drive causes an
increase in iso-orientation activity (the ``surround'' response is greater
than ``blank''), while ``surround'' stimulation in the presence of high
contrast ``center'' stimulation causes a decrease in
activity (the ``full-field'' response is less than the ``center'' response).
Data from a representative animal (different from the one shown in figure 5 but with similar activity maps) are shown in figure 6. A high contrast ``surround'' stimulus caused an increase in cortical activation compared to a ``blank'' stimulus. However, a ``full-field'' stimulus (i.e., a high contrast center plus surround) caused a reduction in activity compared to a high contrast ``center'' stimulus alone. This biphasic modulation of signal strength compares well with the modulation of neuronal activity by these same stimuli calculated from the model. The suppression of activity by the ``full-field'' stimulus is even greater when the ``full-field'' response magnitude is compared to the sum of ``center'' and ``surround'' response magnitudes (figure 6). The amount of modulation depends on several factors, including the contrast of the ``center'' and ``surround'' stimuli (see figure 6) as well as the extent of extraclassical receptive field engaged by the ``surround'' stimulus.