
Next: Computational and Functional
Up: Introduction: The Emerging Understanding
Previous: Introduction
The purpose of this section is to review central results on the anatomy
and development of lateral connections in the cortex. Some of these
results are still controversial, and the details will undoubtedly be
refined as more experimental results are obtained. However, a fairly
good understanding of the area already exists and can serve as a basis
for computational modeling.
The essential properties of long-range lateral connections are outlined
in the Computational and ... section. Although most of the data comes from the low-level visual areas,
it is important to note that patterned lateral connections are not just
a feature of the primary visual cortex: They are found at all levels of
the cortical hierarchy and appear to be a general feature of cortical
organization [2].
- Extent:
- Long-range lateral connections form a dense network
within the cortex. Each connection extends over several millimeters and
gives rise to clusters of axon endings at regular intervals
([12,14,37];
figure 1). The higher the area, the longer the range of
lateral connections. For example in the primary visual cortex, where the
receptive fields of neurons are very small and local, the average extent
of long-range connections is approximately 1.5 mm, or 10% of the length
of the cortical surface. In contrast in the inferotemporal cortex (area
7a), which is one of the highest visual centers and has large receptive
fields, these connections can range up to 10 mm, and the average extent
ranges between 44% and 90% of the cortical length
[2]. Lateral connections appear to be recurrent, that
is, if area A connects to B, then B connects back to A, although the
evidence on this aspect is rather fragmentary at present.
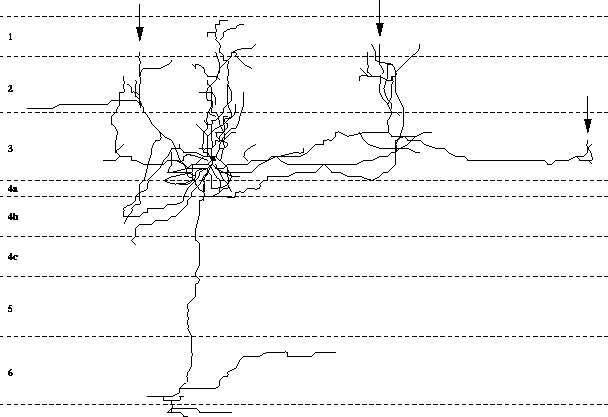
Figure 1: Long-range
lateral connections of a typical neuron in the visual cortex. Lateral
connections, also sometimes called horizontal or intrinsic connections,
run parallel to the cortical surface. In the visual cortex they run over
distances covering several degrees of the visual field, and sprout
branches at intervals (marked by arrows). The branches form a local
cluster of connections to other cells in the region. Such clusters occur
only in regions with similar functional properties as the parent
cell. Adapted from [15].
- Synapses:
- The size of the dendritic arbor of pyramidal cells in
layers 2 and 3 matches the axonal patches made by each horizontal
connection in a cluster. Horizontal connection patterns in different
layers (2, 3, 5, and 6) are aligned. The long-range horizontal
connections have primarily excitatory synapses. About 70-80% synapse on
excitatory pyramidal cells, while the remaining 20-30% seem to synapse
on inhibitory interneurons [13,33].
Imaging studies and other measurements indicate a substantial amount of
long-range inhibition in the cortex, more than predicted by the above
80-20 rule [17,18,19].
- Patterns:
- Long-range connections are clustered in patches whose
distribution corresponds closely to the organization of receptive fields
in the sensory map. For example in the mature visual cortex, lateral
connections primarily run between areas with similar receptive field
properties, such as iso-orientation columns
[13,15]. The connections are elongated
along the orientation axis of each neuron. In the immediate vicinity of
each neuron, the patterns are relatively unspecific to orientation, but
over larger distances, they are closely matched to orientation
preference. To a lesser degree, the patterns are also shaped by other
perceptual features such as ocular dominance and spatial frequency
[3,9,27,28,43].
Under special conditions such as strabismus (squint-eye), the dependency
on ocular dominance can become quite strong. In the strabismic
cat, long-range connections run between ocular dominance columns, and
their clustering is not influenced by orientation column patterns.
- Development:
-
Immediately after birth, lateral connections grow exuberantly and to
long distances in a short period. In the coming weeks, they get slowly
pruned into well-defined clusters
[7,8,26,29].
What process drives such pruning? Enormous amounts of
genetic information would be required to specify each connection and
each synaptic weight of the neurons in a cortical map. Instead, it
appears that lateral connections develop in an activity-dependent
manner. Several observations support this view:
- When the primary visual cortex (of the cat) is deprived of visual
input during early development, lateral connectivity remains crude and
unrefined [8].
- The patterns of lateral connection clusters can be altered by
changing the input to the developing cortex. The resulting patterns
reflect correlations in the input. For example, when a kitten is made
strabismic, thereby removing correlations between the
visual inputs in the two eyes, the lateral connections in the primary
visual cortex organize differently, linking only the regions responding
to the same eye [28].
- In the mouse somatosensory barrel cortex, sensory deprivation (by
sectioning the input nerve) causes drastic decreases in the extent and
density of lateral connections in comparison to a normally reared animal
[32].
These observations suggest that the development of lateral connections,
like that of afferent connections, depends on cortical activity caused
by external input, and represents correlations in the input. Moreover,
the development of the two kinds of connections appears to be
strongly related. The development of lateral connections happens at
approximately the same time as the afferent connections organize into
topographic maps [6,26]. Although
each individual lateral connection is weak, their total effect on neural
activity can be substantial [13], and thereby affect
the development of afferent connections. Changes in afferent
connections then change the activity patterns on the cortex, which in
turn influences the organization of lateral connections. The development
of both sets of connections thus appears to proceed synergetically and
simultaneously, eventually evolving to a state of equilibrium in the
adult animal.
Some of the above findings are still preliminary and will require
further confirmation and more detailed quantification. However, several
questions about the computational mechanisms and functional role of
lateral connections can already be addressed, as will be discussed below.
Next: Computational and Functional
Up: Introduction: The Emerging Understanding
Previous: Introduction