


Figure 1: Steerable and
shiftable filters (receptive fields). The properties of
steerability and shiftability allow a continuum of orientation
preferences to be obtained from a small number of discretely tuned
orientation-selective basis RFs
(the Shaping the RFs section).
Several models for the formation of the original set of discrete
orientations by projections from the lateral geniculate nucleus (LGN)
to the striate cortex have been offered in the past
[18,47]. In particular, it has been argued that the
receptive fields at the output of the LGN are already broadly tuned
for a small number of discrete orientations (possibly just horizontal
and vertical), and that at the cortical level the entire spectrum of
orientations is generated from the discrete set present in the
geniculate projection
[47]. In the present model, the size
of the original discrete orientation columns is determined by the
minimal cortical separation of cells with non-overlapping RFs, called
the point image
[24]. Thus, the model incorporates a
network of orientation columns, whose size is determined by the
diameter of their constituent RFs. Each column is tuned to a specific
angle, and located at an approximately constant distance from another
column with the same orientation tuning. The RFs of adjacent units
with the same orientation preference are overlapping, with the amount
of overlap determined by the number of RFs built into the network.
The preferred orientations are distributed uniformly in the range
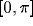 . Each RF is modeled by a product of a 2D Gaussian
. Each RF is modeled by a product of a 2D Gaussian 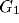 ,
with center at
,
with center at 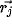 , and an orientation selective filter
, and an orientation selective filter 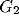 ,
with optimal angle
,
with optimal angle 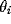 :
: 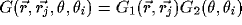 . This model for a
receptive field is equivalent to a directional derivative of a 2D
Gaussian (cf.
[50]).
. This model for a
receptive field is equivalent to a directional derivative of a 2D
Gaussian (cf.
[50]).
According to the theory of shiftable/steerable filters
[14,40], an RF located at
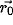 and tuned to the orientation
and tuned to the orientation 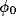 can be obtained by a
linear combination of basis RFs, as illustrated in
Figure 1. The numbers
can be obtained by a
linear combination of basis RFs, as illustrated in
Figure 1. The numbers
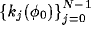 and
and
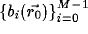 denote the steering and
shifting coefficients, respectively; because orientation and
localization are independent parameters, the
denote the steering and
shifting coefficients, respectively; because orientation and
localization are independent parameters, the 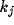 's can be calculated
separately from the
's can be calculated
separately from the 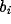 's. The number of steering coefficients
depends on the polar Fourier bandwidth of the basis RF, while the
number of steering filters is inversely proportional to the basis RF
size. Details regarding this scheme and its performance may be found
in
[17].
's. The number of steering coefficients
depends on the polar Fourier bandwidth of the basis RF, while the
number of steering filters is inversely proportional to the basis RF
size. Details regarding this scheme and its performance may be found
in
[17].
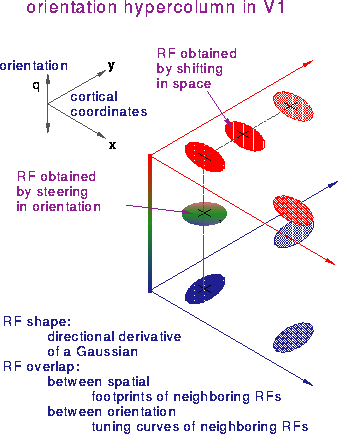
Figure 2: Lateral connections in V1. The
construction of the observed wide variety of orientation tuning
curves from steerable/shiftable basis RFs may be implemented by
the lateral connections in the primary visual area.
The mathematical properties of shiftable/steerable filters outlined
above suggest that the columnar architecture in V1 provides a basis
for creating a continuum of RF properties. Computationally, this
requires that the input to a V1 neuron be a linear combination of
outputs of several RFs, as in Figure 1. Is this
assumption warranted by anatomical and physiological data regarding
cortical interconnection patterns, and, in particular, patterns of
lateral connections? Horseradish peroxidase (HRP) labeling studies
[32] have shown that lateral connections of orientation
columns extend to a range of 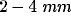 . In other studies that used 2DG
autoradiography and retrograde labeling, connectivity patterns were
superimposed on functional maps
[16]. The results showed
that cells tended to connect to cells of like orientation preference.
The relationship between functionally defined columns and patchy
connections was studied by
[26]. They used optical
imaging techniques to construct functional maps of orientation
columns, then targeted injections of biocytin tracer to selected
functional domains. Their results show that long-range connections,
extending
. In other studies that used 2DG
autoradiography and retrograde labeling, connectivity patterns were
superimposed on functional maps
[16]. The results showed
that cells tended to connect to cells of like orientation preference.
The relationship between functionally defined columns and patchy
connections was studied by
[26]. They used optical
imaging techniques to construct functional maps of orientation
columns, then targeted injections of biocytin tracer to selected
functional domains. Their results show that long-range connections,
extending 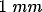 or more, tend to link cells with like orientation
preference. In the short range, up to
or more, tend to link cells with like orientation
preference. In the short range, up to 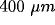 from the injection
site, connections were made to cells of diverse orientation
preferences. The selectivity of the short-range connections is
markedly disrupted, probably because dendritic arbors and axonal
connections freely cross orientation column borders
[25].
from the injection
site, connections were made to cells of diverse orientation
preferences. The selectivity of the short-range connections is
markedly disrupted, probably because dendritic arbors and axonal
connections freely cross orientation column borders
[25].
These findings suggest that the long-range connections, which link
cells of like orientation preference, may provide the inputs necessary
to shift the position of the desired RF, while the short-range
connections, linking cells of diverse orientation preference, may
provide the substrate for steering the RF to an arbitrary angle
(Figure 2). Note that the model requires both excitatory
and inhibitory connections; biological data suggest that this
requirement is not unreasonable. According to
[16], the
majority of long range horizontal connections are excitatory and link
pyramidal cells. Cross-correlation studies
[45] support this
observation. Inhibitory connections come from two sources: first, a
small proportion of postsynaptic cells (as high as 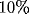 ) may be
inhibitory interneurons; second, it is possible that
orientation-biased cells within cytochrome oxidase-rich blobs in
primates (where high GABA-decarboxylase activity indicates probable
presence of inhibitory synapses) provide inhibitory inputs to the
sharply tuned orientation selective cells
[48].
) may be
inhibitory interneurons; second, it is possible that
orientation-biased cells within cytochrome oxidase-rich blobs in
primates (where high GABA-decarboxylase activity indicates probable
presence of inhibitory synapses) provide inhibitory inputs to the
sharply tuned orientation selective cells
[48].