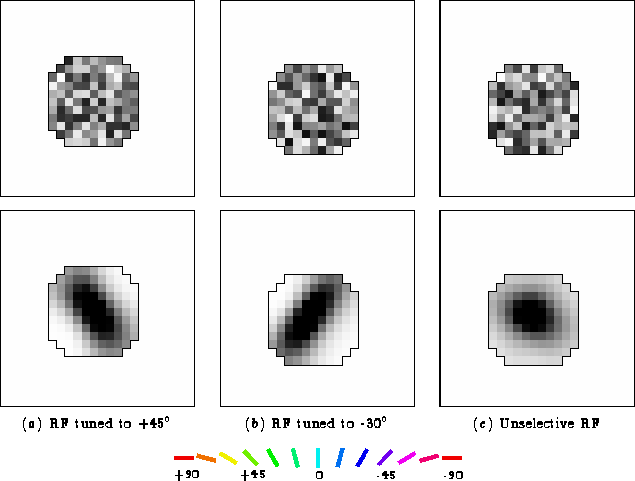
To show how the orientation preferences are located across the network, an orientation map was computed by labeling each neuron with its orientation preference, as determined from its afferent weights. The afferent weights were fitted to an ellipsoidal Gaussian (equation 4.1) using the nonlinear programming package NPSOL (Gill et al., 1986). The orientation of the fitted function was taken to be the orientation preference of that neuron; it is the orientation of the Gaussian that maximally excites the afferent weights. Because the receptive fields were themselves developed based on Gaussian inputs, their orientation preferences were generally unambiguous, and other methods of computing the orientation map would show very similar results.
Figure 4.3 shows the global organization of the
receptive fields in the network before and after training.
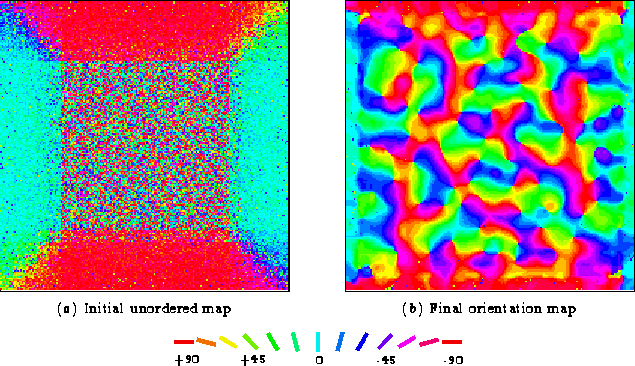 |
![[*]](foot_motif.gif) The results suggest that Hebbian self-organization of afferent
weights, based on recurrent lateral interactions, could underlie the
development of orientation maps in the cortex.
The results suggest that Hebbian self-organization of afferent
weights, based on recurrent lateral interactions, could underlie the
development of orientation maps in the cortex.