A Model of Visually Guided Plasticity of the Auditory Spatial Map in the Barn Owl
Abstract
In the barn owl, the self-organization of the auditory map of space in the
external nucleus of the inferior colliculus (ICx) is strongly influenced
by vision, but the nature of this interaction is unknown.
In this paper a biologically plausible and minimalistic model of ICx
self-organization
is proposed where the ICx receives a learn
signal based on the owl's visual attention.
When the visual attention is focused in the same spatial location
as the auditory input, the learn signal is turned on, and the map
is allowed to adapt.
A two-dimensional Kohonen map is used to model the ICx, and
simulations were performed to evaluate how the learn signal would affect
the auditory map. When primary area of visual attention was shifted at
different spatial locations, the auditory map shifted
to the corresponding location. The shift was complete when done early in
the development and partial when done later. Similar results have been
observed in the barn owl with its visual field modified with prisms.
Therefore, the simulations suggest
that a learn signal, based on visual attention,
is a possible explanation for the auditory plasticity.
Introduction
In the brain, several computational maps process
sensory information. The maps transform the sensory input into
a localized activity on the map, which can be easily accessed
by other neural processes. These maps self-organize so that the
input space is represented topologically on the map
(Knudsen et al., 1987). Typically each map is concerned with only one sensory modality.
However, the barn owl is unusual in that its auditory map
is strongly influenced by a different modality,
vision.
Some kind of visual influence on the auditory map is known to exist because the
auditory map adapts even when only vision is distorted.
The auditory map in the external nucleus of the inferior colliculus (ICX)
projects the auditory input to the
optic tectum. In the optic tectum, the auditory information is combined with
visual input to form a bimodal topographic map of space. This spatial map
allows the barn
owl to locate its prey using either visual or auditory cues.
In order to support the bimodal organization in the optic tectum, the auditory
map in the inferior colliculus must be structured appropriately;
a vision-based calibration signal must be involved in its self-organizing process.
In this paper, the
hypothesis that a learn signal, based on coincidence of visual attention
and auditory input, mediates the auditory plasticity of the ICx, is proposed
and evaluated
computationally.
Auditory and Visual Localization in the Barn Owl
To motivate discussion of the learn signal, let us first review the
roles the inferior colliculus and the optic tectum have on localization.
The barn owl primarily uses sounds to localize its
prey in the dark, and it can do this with an accuracy surpassing that of
most birds and mammals (Payne, 1971). The auditory maps that give the barn owl its
extraordinary abilities are located in the inferior colliculus.
There are three subdivisions of the inferior colliculus known as the central nucleus
(ICc), external nucleus (ICx), and superficial nucleus (ICs). The ICs is a relatively
small portion of the inferior colliculus, and little is known about its function other
than there may be a few projections to the ICx (Knudsen, 1983).
The neurons in the ICc, however, are sharply tuned to frequency and
tonotopically organized, and the neurons
in the ICx are broadly tuned to frequency and spatiotopically organized. The ICx
receives its input from the ICc.
The frequency-coded auditory input to the ICc is transformed to spatial location in the
ICx by the projection from the ICc to the ICx. This transformation process, involving
interaural level differences (ILD) and interaural time differences
(ITD), can unambiguously determine the location, in azimuth and elevation, of the
source of the auditory input (Brainard et al., 1992; Knudsen, 1987).
The spatiotopic organization that exists in the ICx is projected to the optic tectum.
In the optic tectum (OT), there is a general bimodal map of space that
responds to both visual
and auditory input (Knudsen, 1982).
This map enables the owl to determine the location
of its prey either using visual cues or auditory cues.
Experiments with visual stimuli
have shown that neurons in the optic tectum are organized according to
azimuth and elevation, and a visual stimulus in a certain area of space
causes the neurons representing that area of space to fire much more
rapidly (Knudsen & Konishi, 1978; Knudsen, 1982). The map
responds to auditory stimuli in the same way. Most of
the neurons that respond to visual stimuli also respond
to auditory stimuli. In addition,
the location of an auditory response is usually very close to the location of the
corresponding visual response. This way, both modalities lead to similar
responses, and other neural structures can process location information
in the same way, irrespective of the originating modality.
Since the map in the OT is formed by merging two different modalities,
auditory and visual input must have a compatible structure
in order to assure that the bimodal map will encode the same
location for both inputs.
Furthermore, since the region of the bimodal map that corresponds to
the area directly in front of the owl is magnified (occupying a disproportionately
large portion of the map), the auditory spatial map in the ICx
must have the same amount of magnification in this area in order for the
visual and auditory locations to correspond. The structure of the retina
causes magnification of the visual input. Since there is no corresponding
mechanism that magnifies auditory input, the auditory map in the ICx must be conforming
to the visual map in the optic tectum. Therefore, it seems that a visually
based calibration signal must exists which guides the development of the
auditory map in the ICx.
Several experiments have been performed to determine what influence
vision and hearing have on the formation of the bimodal map in the optic
tectum
(Knudsen, 1985, 1988; Knudsen & Brainard, 1991; Knudsen & Knudsen,
1985a, 1985b, 1990).
Since the OT receives its auditory input from the ICx, any changes in
the representation of auditory space in the OT reflect the plasticity that is
occurring in the ICx due to vision. For example,
prisms or occluders were mounted over the owl's eyes to manipulate
the visual information the owl received (Knudsen & Knudsen, 1985a).
The adaptation usually took
weeks and the prisms were left on for a period of months. While the
owls still had the prisms on, their auditory localization abilities
were tested by having the owl orient its
head directly at the location of an auditory stimulus.
However an owl, wearing right shifting prisms, localized to the right of the
auditory stimulus. Even though the owl received correct auditory information,
it could not accurately locate the stimulus; instead the owl chose a location
that conformed to the visual distortion created by the prisms.
This is an instance where a shifted visual signal causes the formation of an
abnormal auditory spatial map in the ICx, and therefore in the OT, even though there
were no distortions in the auditory input.
Vision is used as a recalibration mechanism for the auditory spatial map in the
ICx, even if the visual cues are incorrect. These experiments show an innate
dominance of vision over audition.
Where does the visual recalibration signal come from? The ICx does not respond to any
visual inputs, so there are no direct visual signals available for comparison
at the ICx. Anterograde labeling revealed that there
was no direct
feedback from the optic tectum to the ICx either (Knudsen & Knudsen, 1983).
Previous computational models of visual calibration in the ICx map
have relied on such connections, modeled by backpropagation of an error signal and/or
a reinforcement signal (Rosen et al., 1994; Pouget et al., 1995). In addition,
these models did not address
how the two-dimensional maps in the ICx could self-organize from
the visual input.
It has been confirmed
that the synaptic changes that alter the auditory maps occur in the ICx itself,
and not in the lower centers or in the optic tectum (Brainard & Knudsen, 1993).
To date, the nature
of the recalibrating signal to the ICx is not well understood.
In the remainder of this paper, a simple biologically plausible mechanism for
the self-organization and plasticity of the ICx is proposed. Simulations are
performed to demonstrate the plasticity of the ICx and the effects of the
proposed learn signal. The results are then discussed along with some possible
future areas of research.
The Learn Signal Model
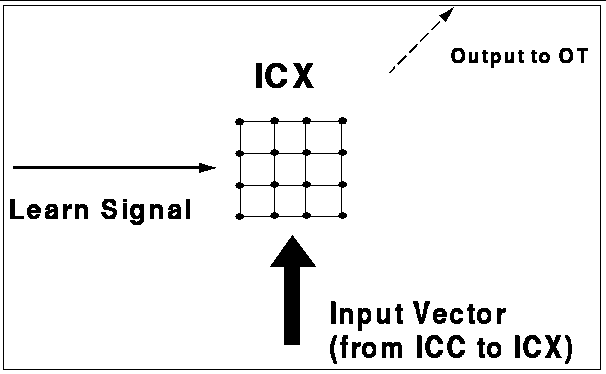
Figure 1: The ICx model. The two-dimensional feature map stands for the auditory spatial
map in the ICx of the barn owl. The input from the ICc
is a vector that is propagated
to each node in the network. The learn signal is either on
or off, and determines whether the map will be adapted. The neurons of the
ICx project to the optic tectum.
The model is based
on the self-organizing feature map (Kohonen, 1981, 1989, 1990),
which is an abstraction
of the biological mechanisms that give rise to topographic maps.
Here, a two-dimensional Kohonen map models the auditory spatial map in
the ICx. The spatial location is assumed to be computed by the
projection from the ICc to the ICx, and the map network receives the
resulting spatial representation vector as its input (figure 1).
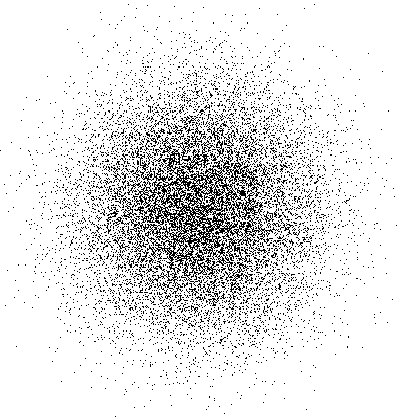
These auditory input vectors are uniformly distributed
since a sound can originate at any location in space (figure 2a).
 |
 |
 |
| (a) Auditory input space |
(b) Visual attention centers |
(c) Auditory spatial map |
Figure2: Self-organization
of the normal topographic ICx map.The square region in (a) is a
two-dimensional representation of
the auditory space, and
the inputs are uniformly distributed in this space.
Figure (b) shows the distribution of visual attention centers
that were used to determine the on/off value of the visual instructive signal.
The attention is distributed about the center of the input space in
a gaussian fashion, so that the center is attended to more frequently than
the periphery. When trained with these signals, a topographic
map of the input space develops, as shown in figure (c). The width
of the map corresponds to the spread of the attention signal.
How could visual input calibrate the auditory map formed by the Kohonen
algorithm? Because the ICx does not respond to any visual inputs directly,
the calibration signal, while
visually based, must be of a different form. A simple learn signal
that turns the synaptic learning on or off is proposed in this paper. When the visual
attention and the location of the sound source coincide, the
learn signal is turned on and allows the map to adapt. Thus, the
signal forces the map to learn the portion of the input space
currently attended to.
Since the owl attends more often to the center of its visual field than to
the periphery, a gaussian distribution around the center of the
input space is used to generate the visual attention (figure 2b).
In the Kohonen map the neuron that is most
similar to the input vector is known as the excitation center. The excitation
center of the auditory map, for input v is defined as the neuron r'
for which
Forall r: ||v - w(r)|| < =
||v - w(r')||, (1)
where r are the nodes in the network and w(r) is the vector of
weights.
The excitation center is the image of the auditory input on the map.
During training the learn signal must be computed. If the Euclidean distance
between the auditory input
and the visual attention is
within a certain threshold theta, the learn signal will be on; otherwise the signal
is off. When the signal is on, the synaptic strengths of the neighborhood
around the excitation center are modified according to the standard feature map
learning algorithm
w'(r) = w(r) + a(v = w(r)), (2)
where a is the learning rate. Adaptation occurs only when
the owl is attending to the area of space where the sound originates, that is,
when the auditory input and visual attention coincide.
Simulations
 |
 |
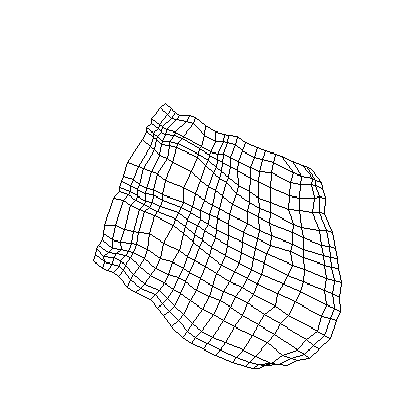 |
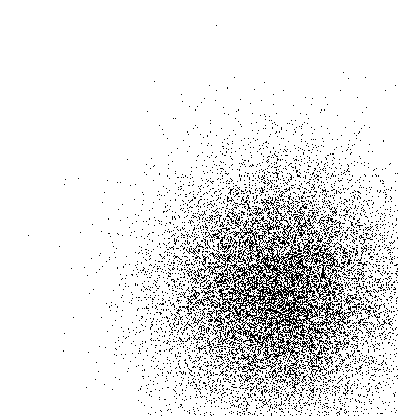
| (a) Shifted visual attention centers |
(b) Attention shifted from start |
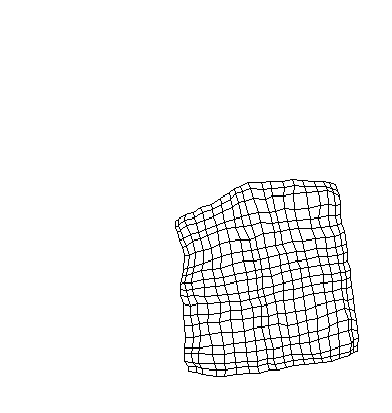
(c) Attention shifted after some training |
Figure 3: Self-organization
of the ICx map with shifted inputs.
Figure (a) shows the shifted distribution of visual attention centers.
Figure (b) displays the map trained from the start with the shifted attention
signal. The entire map has shifted to coincide with the position of the
attention signal. However, if the attention signal is shifted midway
during training, only the portion of the map close to the new attention
center shifts, as shown in figure (c).
Simulations were performed using a 20x20 neuron network
with random initial values for all weights.
Each training trial consisted of four steps: (1) An input vector was generated and the excitation
center was determined using equation 1.
(2) An attention center was generated and (3)
compared with the excitation center. If the signals were relatively close
(within the threshold theta, the learn signal was turned on;
otherwise the learn signal was off. (4) If the learn signal was on, synapses
were modified according to equation 2. A total of 20,000 training trials
were required for the map to organize.
A series of experiments were performed to simulate the different experimental
conditions on owls with and without prisms. The first experiment simulated
the control case where the owl is allowed
normal vision. The visual attention is generated from a gaussian that
is centered over the input space (figure 2 b).
The resulting topographic
map is shown in figure 2c. The map
is centered in the input space, and the extent to which the map
covers the space is determined by the spread of the gaussian
attention signal. Thus, the learn signal focuses the map to the
most attended portion of the input space.
To simulate the development of the
ICx map with prisms, the center of the gaussian distribution of attention
was shifted relative to the input space (figure 3a).
The second experiment simulated the owl wearing the prisms
before its eyes had opened. In this case, the gaussian was shifted
before any training steps.
The resulting network had a similar shape as in the control case,
but the entire
network had shifted in the direction of the learn signal
(figure 3b). Here, the map was forced to learn the
inputs in the shifted region.
In the third experiment, the center
of the gaussian was shifted after 10,000 training trials,
simulating a period of normal development, after which the
prisms were placed over the owl's eyes. Initially the map was forming in the
center of the input space. After the shift occurred, the map slowly
moved towards the new attention center. The area of the map furthest away from the
signal was slower to adapt and looked similar to the map where the learn
signal is centered. In conclusion, if the shift was introduced right from the start,
the network learned only the attended region (figure 3b);
however, if the shift
was introduced in the middle of training, the map shifted only partially
(figure 3c). These results are in agreement with those
observed experimentally in the barn owl (Knudsen & Knudsen, 1990).
Discussion
The proposed model of the visually guided plasticity in the ICx is minimalistic
in its assumptions and
biologically well-motivated. The single Kohonen map represents the auditory
spatial map that exists in the ICx of the barn owl. It is not necessary
to model the OT because the plasticity of the auditory map occurs at the
level of the ICx. The model shows that a simple on/off learn signal, based
on the coincidence of visual attention and auditory input, is a sufficient
explanation for auditory map plasticity. The learn signal does not need
connections from the optic tectum to the ICx, which several other models rely on,
because the learn signal is not based on a comparison of the visual
and auditory inputs in the OT.
The learn signal is also much simpler than an error signal, which would include
information such as the magnitude and location and even direction of error.
The learn signal could originate from the higher cortical areas
of the brain where visual input has already been processed, and the location
for visual attention formed.
In humans and other animals, sensory modalities are combined to give
a single cohesive view of the world. In certain cases the perception of
the world can be distorted, as in an illusion, because of conflicting
information from different sensory inputs. The visual dominance in the
formation of the multimodal map in the barn owl gives insight into the
mechanisms used for the integration of different sensory modalities and
how one modality can distort the perceptions of other modalities.
In future work, we plan to extend the model with more realistic neurons
with lateral connections where the weight modification
process is completely unsupervised (e.g. Sirosh & Miikkulainen, 1994).
Furthermore, we plan to include the ICc to ICx connections, which are
responsible for computing the spatial input representation from the
frequency-specific
interaural level differences (ILD) and interaural time differences (ITD).
This way the ICx would be organized according to direct inputs from
the ICc instead of intermediate spatial representations as in the current
model. The bimodal map that exists in the OT could also be included.
This map would not only represent the visual input,
but it would also incorporate the auditory input from the Kohonen map
in the existing model. Such a comprehensive model would be a major
step toward verifying that the learn signal is still sufficient for
the plasticity of the ICx on this large scale, and that it causes
a similarity in structure of both the auditory and visual spatial maps so that
the merging done in the OT is possible.
Conclusion
The simulations reported in this paper demonstrate that a simple
visually-based learn signal is a sufficient explanation of
the auditory plasticity observed in the ICx of the barn owl.
Unlike in previous models, an error signal is not necessary
to calibrate the auditory map. Rather, the simple
coincidence of visual attention and spatial location of auditory input
may alone drive the plasticity of the ICx. The coincidence signal may be
generated in the cortical area that is responsible for attention.
Direct feedback projections from the optic tectum or close coupling
of the OT and ICx are not necessary. In the future, biological experiments
should be performed to verify whether such a learn signal exists, and
also to determine the signal pathway to the ICx from the higher cortical
areas, possibly via the ICs.
References
Brainard, M.S. & Knudsen E.I. (1993). Experience-dependent
plasticity in the inferior colliculus: A site for visual calibration
of the neural representation of auditory space in the barn owl.
The Journal of Neuroscience 13, 4589--4608.
Brainard, M.S., Knudsen, E.I. & Esterly S.D. (1992). Neural
derivation of sound source location: Resolution of spatial ambiguities
in binaural cues. The Journal of the Acoustical Society of
America 91, 1015--1027.
Knudsen, E.I. (1982). Auditory and visual maps of space in the
optic tectum of the owl. The Journal of Neuroscience 2, 1177--1194.
Knudsen, E.I. (1983). Subdivisions of the inferior colliculus in
the barn owl (tyto alba). The Journal of Comparative
Neurology 218, 174--186.
Knudsen, E.I. (1985). Experience alters the spatial tuning of
auditory units in the optic tectum during a sensitive period in the barn
owl. The Journal of Neuroscience 5, 3094--3109.
Knudsen, E.I. (1987). Neural derivation of sound source location
in the barn owl. An example of a computational map. The Annals of the
New York Academy of Science 5, 3094--3109.
Knudsen, E.I. (1988). Early blindness results in a degraded auditory
map of space in the optic tectum of the barn owl. The Proceedings of
the National Academy of Science 85, 6211--6214.
Knudsen, E.I. & Brainard, M.S. (1991). Visual instruction of the
neural map of auditory space in the developing optic tectum. Science
253, 85--87.
Knudsen, E.I., du Lac, S. & Esterly, S.D. (1987). Computational
maps in the brain. Annual Review of Neuroscience 10, 41--65.
Knudsen E.I. & Knudsen P.F. (1983). Space-mapped auditory projections
from the inferior colliculus to the optic tectum in the barn owl
(tyto alba). The Journal of Comparative Neurology
218, 187--196.
Knudsen, E.I. & Knudsen P.F. (1985a). Vision calibrates sound localization
in developing barn owls. The Journal of Neuroscience 9(9), 3306--3313.
Knudsen, E.I. & Knudsen P.F. (1985b). Vision guides the adjustment of
auditory localization in young barn owls. Science 230, 545--548.
Knudsen, E.I. & Knudsen P.F. (1990). Sensitive and critical periods
for visual calibration of sound localization by barn owls. The Journal of
Neuroscience 10, 222--232.
Knudsen, E.I. & Konishi, M. (1978). A neural map of auditory space in
the owl. Science 200, 795-797.
Kohonen, T. (1981). Automatic formation of topological maps of patterns
in a self-organizing system. In Proceedings of the 2nd Scandinavian Conference
on Image Analysis (pp. 214--222). Espoo, Finland: Pattern Recognition Society of
Finland.
Kohonen, T. (1989). Self-Organization and Associative Memory, chapter 5.
Berlin; Heidelberg; New York: Springer.
Kohonen, T. (1990). The self-organizing map. Proceedings of the IEEE
78, 1464--1480.
Payne, R.S. (1971) Acoustic location of prey by barn owls (tyto
alba).
The Journal of Experimental Biology 54, 535--573.
Pouget, A., Deffayet, C. & Sejnowski, T.J. (1995). Reinforcement learning
predicts the site of plasticity for auditory remapping in the barn owl. In
Advances in Neural Information Processing Systems 7 San Mateo, CA: Morgan
Kaufman Publishers.
Rosen, D.J., Rumelhart, D.E. & Knudsen, E.I. (1994). A connectionist
model of the owl's sound localization system. Advances in Neural Information
Processing Systems, 6.
Sirosh, J. & Miikkulainen, R. (1994).
Cooperative self-organization of afferent and lateral connections in
cortical maps. Biological Cybernetics
71(1), 66--78.